 Struttura generale:
Struttura generale:
Pteropus medium
Struttura generale:
La struttura in generale presenta tipici adattamenti al volo riscontrabili anche negli uccelli, quali ossa alleggerite negli spessori dellosso compatto delle diafisi delle ossa lunghe, arti superiori modificati ad ali per permettere il volo. A differenza di quanto si vede negli uccelli, i chirotteri hanno mantenuto invariato il numero delle dita che vengono ricoperte dal patagio (membrana cutanea con la funzione di aumentare la superficie totale del corpo al fine di acrescere la portanza durante il volo). Tra le caratteristiche di questordine di mammiferi notiamo la presenza delle clavicole (probabilmente questo fu uno dei caratteri che ispirarono gli antichi ad accumunare nella classificazione i chirotteri ad i primati), che aumentano la mobilità dellarto toracico, uninversione dellestremità dellarto pelvico che favorisce il meccanismo di prensione degli arti per la particolare locomozione e stazione dei Chirotteri. Conviene comunque analizzare le singole parti scheletriche per poter più facilmente riconoscere gli adattamenti propri di questo ordine di animali ed in particolare della famiglia degli Pteropus. Si prenderanno in esame prima lo scheletro postcraniale nelle sue varie parti e in fine il cranio suddivisibile virtualmente in: il cranio propriamente detto, mandibola e, in fine, dentatura. In ognuna di queste parti scheletriche si ritrovano importanti punti di riferimento per il riconoscimento del genere, per la comprensione di moltissimi adattamenti ecologici dellanimale e per un inquadramento dellordine nel contesto degli altri mammiferi.
Lo scheletro postcraniale è suddivisibile a sua volta in varie parti analizzabili inizialmente singolarmente, in seguito nel contesto globale del sistema scheletrico dellanimale: esse sono il rachide, la gabbia toracica formata da sterno e coste, i cinti pelvico e toracico ed in fine gli arti toracico e pelvico.
Rachide
Si suddivide in tratto cervicale, toracico, lombare, sacrale e coccigeo. Il numero delle vertebre per ogni segmento è: sette per il cervicale, tredici per il toracico, cinque per il lombare, quattro per il sacrale e quattro per il coccigeo; trentatré vertebre in tutto. Il tratto cervicale si presenta, nellinsieme piuttosto allungato; le singole vertebre sono piuttosto robuste, in particolare modo latlante e lepistrofeo. I processi trasversi, bifidi, sono molto pronunciati; evidentemente lo sviluppo dei muscoli del collo è notevole. Rapportato alle altre porzioni del rachide, questo tratto è più massiccio. I processi spinosi sono pochissimo sviluppati o assenti per tutto il tratto cervicale e toracico; a livello lombare sino al tratto coccigeo compreso appaiono più rilevati. I muscoli dorsali sono molto sviluppati a livello del treno posteriore rispetto a quello anteriore. Le vertebre coccigee si presentano piuttosto compresse medio lateralmente e lultima vertebra si articola saldamente con i due ischi; per cui non supra il margine caudale del bacino.
Gabbia toracica
Le coste sono in numero di tredici coppie, di cui sei sternali. La loro forma e struttura è molto particolare e, nellambito dei mammiferi, direi unica. La forma si presenta piuttosto allargata nel tratto prossimale; ad un terzo, circa, della lunghezza delle coste si riduce bruscamente è prosegue praticamente con forma cilindrica. Le coste sternali no presentano cartilagini costali per il collegamento con lo sterno. Bensì esse sono sostituite da formazioni ossificate che si articolano prossimamente con le coste e distalmente con lo sterno. La prima costa si presenta di gran lunga più larga e robusta delle altre, come pure la cartilagine costale ossificata è di forma pressoché quadrangolare. Questa particolare configurazione della gabbia toracica è molto simile a quanto è osservabile negli uccelli, e questo è comprensibile in quanto entrambi questi gruppi di animali sono adattati al volo e di conseguenza è comprensibile anche una certa convergenza adattativa. La struttura così conformata si presenta assai irrigidita è resistente e quindi adatta a sopportare le pressioni che si sviluppano durante il volo. Nelle coste asternali si ritrovano le cartilagini costali come negli altri mammiferi anche se le prime presentano ancora un certo grado di mineralizzazione. Anche lo sterno si presenta conformato per permettere linserzione dei potenti muscoli pettorali che permettono a questi animali di volare. La struttura tende ad assomilgiare a quella degli uccelli anche se con alcune differenze. La prima sternebra, si presenta molto voluminosa e dalla sua linea sagittale sulla superficie ventrale si erigono due grossi e robusti processi che si dispongono tra loro a V. Essi occupano praticamente tutta la superficie della sternebra sulla linea mediana. La seconda sternebra non presenta alcun rilievo; dalla terza alla sesta si ha un processo mediano che decresce procedendo caudalmente. Questa disposizione, similmente a quanto si può rilevare negli uccelli, aumenta enormemente la superficie deputata allinserzione dei muscoli pettorali, anche se a differenza degli uccelli la carena, se così la si può definire, non ha una superficie continua ma è interrotta da due docce.
Cinto toracico
Il cinto toracico è formato, in conformità
con gli altri mammiferi dalla scapola e dalla clavicola; la presenza di
questultima aveva sicuramente contribuito a indurre gli scienziati del
settecento di accumunare i chirotteri ai primati. Questo complesso assume
un ruolo di fondamentale importanza durante il volo.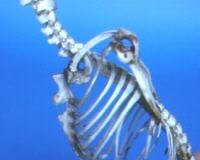
Infatti rappresenta il fulcro sul quale si articola larto toracico responsabile del movimento aereo. La sua struttura è piuttosto particolare: la clavicola si presenta eccezionalmente robusta rispetto agli altri mammiferi, piuttosto allungata in rapporto alla scapola, arquata a concavità caudale. In pratica si dispone parallela al piano sagittale mediano portando la scapola in posizione dorsale. La sezione è appiattita in senso latero mediale e grosso modo assume una forma ovale schiacciata. Notevolmente ampie si presentano le epifisi che portano le superfici articolari per lo sterno e per la scapola. La scapola assume una forma di triangolo isoscele fortemente allungato in senso cranio caudale, il processo coracoideo si presenta notevolmente sviluppato. La spina della scapola divide chiaramente la sua superficie dorsale in una fossa sopraspinata ed in una sottospinata più ampia. Caudalmente la scapola termina appuntita nel vertice dellipotetico triangolo. La cavità glenoidea, molto ampia, si dispone lateralmente e leggermente spostata dorsalmente. Pensando per un attimo al cinto toracico degli uccelli, si notano alcune similitudini, quali ad esempio lesasperazione dellallungamento della scapola in questo gruppo di animali a scapito della larghezza; negli uccelli si ha la presenza di un osso coracoide, che nei pipistrelli è imitato nella sua funzione dal processo coracoideo che è, quindi, molto più sviluppato che negli altri mammiferi. Laspetto complessivo di questo cinto è di grande rigidità, fondamentale per dare stabilità alla struttura corporea durante il volo.
Arto toracico
È forse la porzione scheletrica che maggiormente si è modificata in seguito alladattamento al volo. Fondamentalmente le parti scheletriche sono invariate rispetto agli altri mammiferi ma gli sviluppi delle singole ossa e le proporzioni hanno seguito, ovviamente, levoluzione verso il volo. Per prima cosa bisogna osservare che lo spessore delle ossa è relativamente inferiore a quello che si può constatare negli altri mammiferi in rapporto alle dimensioni, lo spessore dellosso compatto a livello delle diafisi diminuisce sensibilmente. Il motivo di questo adattamento è facilmente intuibile. Si riscontra la medesima cosa negli uccelli, chiaro esempio di confluenza adattativa. È evidente che lalleggerimento della struttura aumenta la capacità di volare. Analizzando l'organizzazione dell'arto, risulta evidente un aumento proporzionale delle ossa responsabili del volo. La lunghezza dellomero, infatti è inferiore a quella delle ossa dellavambraccio; le falangi, in rapporto, sono lunghissime. Lomero non presenta particolari specializzazioni, se non a livello della diafisi distale ove si nota una notevole dimensione della superficie articolare per larticolazione del gomito e lassenza della fossa olecranica; a livello di quella prossimale un notevole sviluppo del trochitere e del trochine per le inserzioni muscolari. Le ossa dellavambraccio, il radio e lulna, assumono una notevole lunghezza, tra i due il predominante, in quanto a dimensione, è il radio, la metà prossimale della diafisi presenta una curvatura a concavità ventro mediale, la rimanente porzione distale ha, invece, un andamento rettilineo. Le epifisi sono notevolmente espanse in senso latero mediale, soprattutto quella distale che partecipa alla formazione dellarticolazione del carpo. Lulna, si presenta estremamente ridotta rispetto al radio, la sua diafisi ha un aspetto filiforme e raggiunge appena i due terzi del radio. Lepifisi prossimale, rispetto agli altri mammiferi, si presenta poco espansa, la sua superficie articolare si presenta pianeggiante, manca il becco dellolecrano e questultimo è molto ridotto. La sua funzione meccanica, a differenza di quanto si piò riscontrare negli altri mammiferi, non comprende il blocco dellestensione dellarticolazione omero radio ulnare (gomito) tramite linserimento del becco dellolecrano nella fossa olecranica ma tende a completare volarmente larticolazione, viene coadiuvata in questa funzione da un osso sesamoideo di una certa dimensione che è compreso nel tendine terminale del muscolo tricipite e posizionato a livello della sua inserzione sullolecrano. La forma e la funzione di questosso sono del tutto simili a quelle riscontrabili nella rotula a livello dellarticolazione femoro tibio rotulea (ginocchio)nei mammiferi. Il carpo è formato da due file di ossa, entrambe formate a loro volta da tre ossa ciascuna; il loro numero è quindi inferiore a quello riscontrabile negli altri mammiferi, dalla forma delle singole parti, sembrerebbe che esse si siano originate dalla fusione di più ossa primordiali. È ragionevole pensare che la struttura del carpo, durante levoluzione dei chirotteri, tendenzialmente abbia seguito la strada della semplificazione rispetto alla condizione primaria più o meno comune nei mammiferi non volatori. Anche se si riduce il numero delle ossa, resta il fatto che questa articolazione continua ad essere molto più complessa rispetto a quella corrispondente degli uccelli. È possibile che i chirotteri, non possedendo le piume e le penne, abbiano bisogno di una maggior mobilità di questa articolazione per poter efficacemente manovrare in fase di volo, considerando anche che abbisognano di manovre spesso molto rapide e strette dato il particolare sistema di orientamento che utilizzano. Il metacarpo, contrariamente a quanto è osservabile negli altri mammiferi, si presenta molto più mobile, è formato da cinque ossa che assumono una lunghezza straordinaria, le diafisi sono molto lunghe e sottili. Le falangi sono due per ogni dito, tranne che per il secondo che ne possiede tre. La prima falange è molto lunga, e a partire dal terzo dito la sua diafisi subisce una torsione in senso ventro laterale in modo che la superficie articolare distale volga lateralmente. La seconda falange del secondo dito, non differisce in forma dalla prima, ma ne è inferiore per quanto riguarda la dimensione, nelle rimanenti dita essa si allunga considerevolmente e assume una forma conica terminando distalmente molto appuntita e sottile.. La terza, nelle prime due dita, assume la forma di un artiglio, più sviluppato nel primo che nel secondo. Questi artigli vengono utilizzati dallanimale per arrampicarsi e muoversi sui rami o sulle rocce. Osservando la forma della superficie articolare prossimale delle falangi terminali delle rimanenti dita, si nota che essa è de tutto simile a quella della seconda falange del secondo dito, per cui penso sia lecito pensare che durante il percorso evolutivo, la terza falange del terzo, quarto e quinto dito sia scomparsa.
Arto pelvico
Per comodità, la descrizione del cranio verrà suddivisa in una prima parte generale in cui si analizzeranno la forma e le proporzioni tra le varie parti ossee, in una seconda parte che riguarderà il "cranio propriamente" detto ossia neuro cranio più splancnocranio con lesclusione della mandibola che verrà trattata separatamente come terza parte e in fine una quarta parte riguardante la dentatura. Generalità sul cranio
Pteropus sp.
Il cranio nel suo complesso si presenta allungato a causa del forte sviluppo del blocco facciale che è anche la causa per la quale volgarmente questi animali vengono chiamati "volpi volanti". La struttura è abbastanza robusta anche se si notano notevoli alleggerimenti in varie parti del teschio. Landamento del profilo dorsale è fortemente incurvato a convessità dorsale. I punto più elevato del cranio è rappresentato dal punto di articolazione delle ossa parietali con le frontali. Partendo dalloccipitale, il profilo dorsale si eleva rapidamente verso lalto sino al punto sopra citato, poi decresce più progressivamente. Questa forma è causata da una rotazione del neurocranio in direzione ventrale facendo perno sulle ossa frontali. Si ottiene quindi, schematizzando, la formazione di un angolo ottuso aperto ventralmente tra il neurocranio e lo splancnocranio. In questo modo i condili occipitali vengono a trovarsi allo stesso livello dellapice dei canini, in una posizione piuttosto ventrale.
Il cranio propriamente detto
Partendo nella descrizione dal neurocranio, si nota che questa porzione è priva di cresta sagittale mediana, le creste nucali sono appena accennate e la protuberanza occipitale esterna è accennata sotto forma di una "bozza". La scatola cranica è piuttosto voluminosa e di forma globosa, la sua superficie è, in generale, liscia tranne che per delle lievi gibbosità sullarea più dorsale. La regione frontale si presenta concava, allungata e stretta, da qui si dipartono due lunghi e appuntiti processi zigomatici che si orientano lateralmente arquandosi ventralmente a delimitare le cavità orbitarie superormente. Alla base di questi processi sono presenti due fori, uno per parte, per il passaggio di vasi. Le arcate zigomatiche si presentano esili, il processo frontale è assente. Il mascellare è abbastanza robusto, sulla sua faccia laerale si notano delle incavature attorno alle rdici dei denti, queste hanno lo scopo di alleggerire il blocco facciale, losso intermascellare ha una forma sub trapezioidale, non si insinua molto tra mascellare e nasale. Contribuisce con le ossa nasali a delimitare rostralmente lapertura nasale che assume una forma a cuore. Alla base del cranio si notano dei processi pterigoidei ben sviluppati e robusti, il palato si presenta di forma rettangolare ed è molto allungato. I processi giugulari sono poco sviluppati e molto appuntiti. Il basiocipitale presenta un notevole sviluppo in relazione alla dimensione totale del cranio.
La mandibola
Le emimandibole che formano la mandibola si uniscono oralmente tramite una sinostosi per cui non si riconosce più larticolazione. La mandibola presenta un margine ventrale orizzontale, il corpo non è particolarmente robusto se non nella sua porzione rostrale che tende ad essere pù spessa della porzione precedente. La branca assume una forma intermedia tra le configurazioni tipiche degli erbivori e quelle dei carnivori; ossia il condilo si trova a metà strada tra la sua posizione ventrale tipica dei carnivori e quella più dorsale degli erbivori. In questo modo si viene a formare un ampio margine osseo che sostituisce il processo angolare e che ha la funzione di ampliare la superficie di inserzione per il muscolo massetere. Si nota, per contro che il processo coronoideo per linserzione del muscolo temporale si presenta ancora molto ampio. Questa particolare situazione porta a pensare che questo genere si sia evoluto da una primordiale situazione di carnivorismo a una volta al erbivorismo, mantenendo ancora caratteri propri di una dieta carnivora, probabilmente è stato vantaggioso mantenere entambe le capacità.
La dentatura
Si compone complessivamente di 38 denti, la formula dentaria
è riassumibile in:
|